
Dynamic processes in live cells
The translation of cellular mRNA to protein is a tightly controlled process often deregulated in diseases such as cancer. Furthering our understanding of the mRNA structural elements, intracellular proteins and signaling pathways that affect protein expression is crucial for the development of new therapies. To this end, we are developing super-resolution microscopy techniques to image and characterize the process of protein translation initiation in live eukaryotic cells. Our custom-made microscope can acquire images at sub-second temporal resolution, allowing us to dissect a variety of dynamic bimolecular processes in live cells.

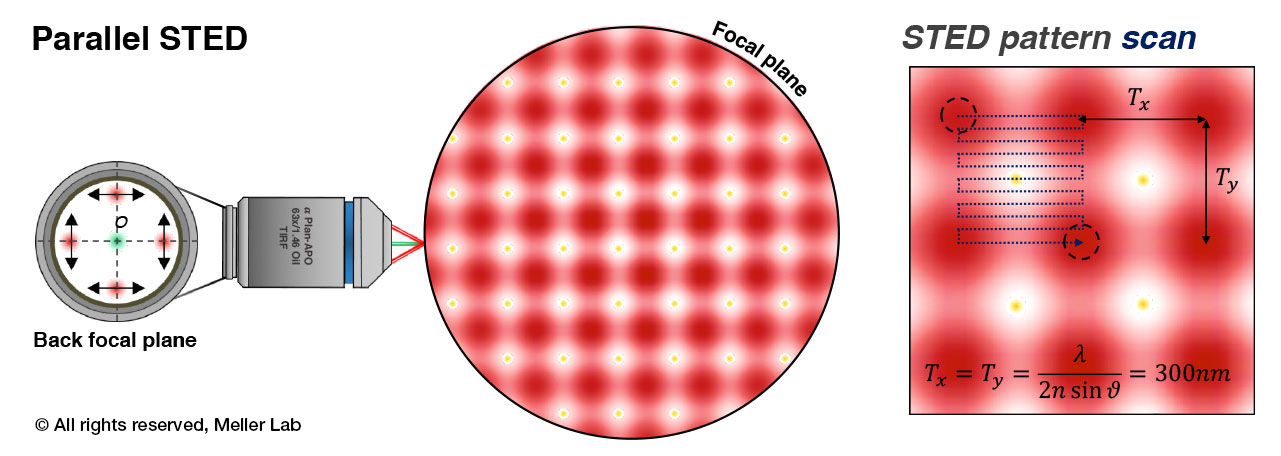
Principles of parallel STED microscopy. An array of STED “cells” is formed by 4-beam interference at the focal plane of the objective lens. This array is quickly scanned with sub-diffraction limit steps to form a super-resolution image with high temporal resolution.
Translation initiation in eukaryotic cells involve the formation of mRNA loops mediated by the interactions of the eukaryotic initiation factors (eIF) and Poly Adenine Binding Proteins (PABP) at the 3’-end of the mRNA. Imaging of the mRNA loop formation in live cells is paramount to understanding the fundamentals of protein synthesis. A fast-scanning parallel STED microscope constructed in our lab enables imaging of these sub-cellular process with sub-second time resolution.

protein localization in Live cells
P53, an important tumor-suppressor gene, has been to shown to preserve genomic stability by recruiting DNA repair proteins upon DNA damage. 53BP1 (p53-binding protein 1) is one such protein that exclusively promotes non-homologous end-joining DNA double strand break (DSB) at damaged chromatin sites. As the dynamics of signaling pathways can be masked when considering a cell population, we have measured the dynamics of 53BP1 in single cells using time lapse fluorescence microscopy. Furthermore, we have exposed MCF10A cells to etoposide, a cytotoxic anticancer drug which prevents re-ligation of DNA strands after unwinding, which invariably results in p53 expression and 53BP1 foci formation several minutes later. As a result, we were able to characterize the time to foci formation, its assembly and subsequent dismantling kinetics.



